 To enhance service speed and avoid tariff delays, we've opened a US warehouse. All US orders ship directly from our US facility.
To enhance service speed and avoid tariff delays, we've opened a US warehouse. All US orders ship directly from our US facility.
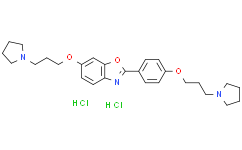




















| Cas No.: | 1345675-25-3 |
| Chemical Name: | E6446 HCl |
| Synonyms: | E6446 dihydrochloride;E6446 (dihydrochloride);BCP29147;AK547246;E-6446 Dihydrochloride |
| SMILES: | Cl[H].Cl[H].O(C1C([H])=C([H])C2=C(C=1[H])OC(C1C([H])=C([H])C(=C([H])C=1[H])OC([H])([H])C([H])([H])C([H])([H])N1C([H])([H])C([H])([H])C([H])([H])C1([H])[H])=N2)C([H])([H])C([H])([H])C([H])([H])N1C([H])([H])C([H])([H])C([H])([H])C1([H])[H] |
| Formula: | C27H37Cl2N3O3 |
| M.Wt: | 522.511 |
| Purity: | >98% |
| Sotrage: | 2 years -20°C Powder, 2 weeks 4°C in DMSO, 6 months -80°C in DMSO |
| Description: | E6446 dihydrochloride is a potent and orally acitve TLR7 and TLR9 antagonist, used in the research of deleterious inflammatory responses. |
| In Vivo: | E6446 (20 mg/kg, p.o.) almost cmlpletely inhibits CpG1668-induced IL-6 production, and dose-dependently suppresses the development of ANA (anti-nuclear antibodies) in mice at 20 and 60 mg/kg[1]. E6446 (20, 60 mg/kg, p.o.) dose-dependently inhibits TLR9 signaling in mice. E6446 (60, 120 mg/kg, p.o.) prevents hyperresponsiveness of TLRs and LPS-induced septic shock in rodent malaria, diminishes TLR responsiveness during acute malaria, suppresses activation of both TLR7 and TLR9[2]. |
| In Vitro: | E6446 dihydrochloride is a potent and orally acitve TLR7 and TLR9 inhibitor. E6446 potently suppresses DNA stimulation of HEK:TLR9 cells, with an IC50 value of 10 nM, but is significantly less effective at suppressing LPS endotoxin stimulation of HEK:TLR4 cells or R848 stimulation of HEK:TLR7 cells. E6446 potently inhibits IL-6 production induced by CpG2216 but is ineffective against induction by the TLR3 ligand poly inosine-cytosine. The ability of E6446 to inhibit TLR7 is ligand dependent, E6446 is a potent inhibitor of IL-6 induction by RNA but a relatively poor inhibitor of IL-6 induction by the small molecule imidazoquinoline ligand R-848. E6446 suppress TLR9-DNA interaction in vitro, with an IC50 in the 1 to 10 µM range[1]. E6446 (0.01-0.03 μM) specifically inhibits TLR9 activation with CpG ODN 2006, and blocks TLR7/8 activated by the imidazoquinoline compound R848 at 2-8 μM. E6446 reduces 50% of TLR4 activation at 30 μM, and shows IC50s of 0.01 μM and 0.23 μM in HEK-TLR9 cells stimulated with oligo 2006 and in human PBMCs stimulated with oligo 2216, respectively[2]. |
| Cell Assay: | E6446 is assayed for the suppression of BALB/c mouse spleen interleukin-6 (IL-6) production in response to stimulation by oligonucleotide CpG1668. E6446 is added to dissociated splenocytes (5 × 105 per well in complete RPMI/10% fetal bovine serum in a 96-well plate) before addition of TLR agonists. Cells are stimulated for 72 hours, and supernatants are removed for ELISA analysis of IL-6. Mouse bone marrow-derived dendritic cells (BMDCs) are generated by culturing BALB/c marrow cells in RPMI containing 100 ng/mL Flt3 ligand for 7 days. Cells (1 × 105) in 50 μL are assayed for IL-6 production after overnight or 24-hour stimulation with various TLR ligands. For studies using human peripheral blood mononuclear cells, Ficoll-separated mononuclear cells are isolated from healthy volunteer donors, washed, and plated with stimulatory oligonucleotide CpG2216 in complete RPMI for 72 hours. Interferon in supernatant is quantified by ELISA[1]. |
| Animal Administration: | Mice[1] MRL/lpr mice are dosed orally five times a week with 20 or 60 mg/kg E6446 or 60 mg/kg hydroxychloroquine beginning at 5 weeks of age. Cytoxan is administered at 50 mg/kg i.p. every 10 days. A serum sample is taken immediately before the beginning of treatment to monitor changes in autoreactive antibodies. Subsequently, serum samples are collected approximately monthly and analyzed for anti-dsDNA by ELISA after 1:500 dilution. Body weights and urine samples are taken at the same interval, and proteinuria is assessed by ChemStrips. Anti-nuclear antibodies (ANA) are assessed using commercially available HEp2 slide kits, with serum diluted to 1:100 in kit buffer. ANA scores are read blinded[1]. |
| References: | [1]. Lamphier M, et al. Novel small molecule inhibitors of TLR7 and TLR9: mechanism of action and efficacy in vivo. Mol Pharmacol. 2014 Mar;85(3):429-40. [2]. Franklin BS, et al. Therapeutical targeting of nucleic acid-sensing Toll-like receptors prevents experimental cerebral malaria. Proc Natl Acad Sci U S A. 2011 Mar 1;108(9):3689-94. |