 To enhance service speed and avoid tariff delays, we've opened a US warehouse. All US orders ship directly from our US facility.
To enhance service speed and avoid tariff delays, we've opened a US warehouse. All US orders ship directly from our US facility.
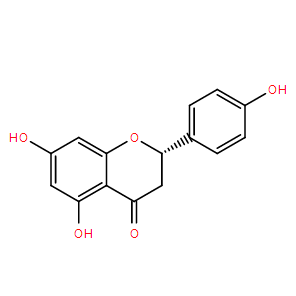




















| Cas No.: | 480-41-1 |
| SMILES: | O=C1C(C(O)=CC(O)=C2)=C2OC(C3=CC=C(O)C=C3)C1 |
| Formula: | C15H12O5 |
| M.Wt: | 272.25 |
| Purity: | >98%, Standard References Grade |
| Sotrage: | 4°C for 1 year, -20°C for more than 2 years |
| Description: | Naringenin is the predominant flavanone in grapefruit; displays strong anti-inflammatory and antioxidant activities. |
| In Vivo: | Naringenin supplementation causes a significant reduction in the amount of total triglyceride and cholesterol in plasma and liver. In addition, naringenin supplementation lowers adiposity and triglyceride contents in parametrial adipose tissue. Naringenin-fed animals show a significant increase in PPARα protein expression in the liver. The expression of CPT-1 and UCP2, known to be regulated by PPARα, is markedly enhanced by naringenin treatment[3]. Naringenin increases hepatic fatty acid oxidation through a PPARγ coactivator 1α/PPARα-mediated transcription program. It prevents sterol regulatory element-binding protein 1c–mediated lipogenesis in both liver and muscle by reducing fasting hyperinsulinemia. Naringenin decreases hepatic cholesterol and cholesterol ester synthesis[4]. Naringenin inhibits TNF-α-induced VSMC proliferation and migration in a dose-dependent manner. Mechanistic study demonstrates that naringenin prevents ERK/MAPK and Akt phosphorylation while left p38 MAPK and JNK unchanged. Naringenin also blocks the increase of ROS generation induced by TNF-α[5]. |
| In Vitro: | Naringenin is shown to inhibit the proliferation of HepG2 cells resulted partly from an accumulation of cells in the G0/G1 and G2/M phase of the cell cycle. Naringenin has been shown to induce apoptosis as evidenced by nuclei damage and increased proportion of apoptotic cells. Naringenin triggers the mitochondrial-mediated apoptosis pathway as shown by an increased ratio of Bax/Bcl-2, subsequent release of cytochrome C, and sequential activation of caspase-3[1]. Naringenin exposure significantly reduces the cell viability of A431 cells with a concomitant increase in nuclear condensation and DNA fragmentation in a dose dependent manner. Cell cycle study shows that naringenin induced cell cycle arrest in G0/G1 phase of cell cycle and caspase-3 analysis reveal a dose dependent increment in caspase-3 activity which leads to cell apoptosis[2]. |
| Cell Assay: | Naringenin is dissolved in DMSO and diluted in cell culture medium. The cells are rinsed with PBS and grown in a medium containing various concentrations of naringenin (50, 100, 150, 200, 250, 300 μM). The solvent DMSO treated cells are served as control. After 24 hrs of treatment, the medium is removed and replaced by another medium containing MTT. Cell viability is measured using the MTT assay[1]. |
| Animal Administration: | Rats: Semi-purified, powdered diets are prepared for concentrations of naringenin: 0, 0.003, 0.006, and 0.012% of diet. After 7 days of acclimatization, rats are assigned to one of four groups, with six animals per group, and fed semi-purified experimental diets for 6 weeks. The experimental diets contain 16% fat, 45.5% sucrose, and different naringenin concentration (0, 0.003, 0.006, or 0.012%) (Table 1). Rats have ad libitum access to food and water during the study period. Food intake and body weight are measured throughout the experiment[3]. Mouse: Eight- to 12-week-old mice are fed ad libitum a rodent standard diet or a high-fat diet containing 42% of calories from fat plus cholesterol (0.05% wt/wt). Naringenin is added to the Western diet at 1 or 3% (wt/wt). Ldlr−/− mice are fed for 4 weeks and C57BL/6J mice for 30 weeks. Food intake is measured daily, and body weight is measured biweekly. Mice are fasted for 6 h before intervention[4]. |
| References: | [1]. Arul D, et al. Naringenin (citrus flavonone) induces growth inhibition, cell cycle arrest and apoptosis in human hepatocellular carcinoma cells. Pathol Oncol Res. 2013 Oct;19(4):763-70. [2]. Ahamad MS, et al. Induction of apoptosis and antiproliferative activity of naringenin in human epidermoid carcinomacell through ROS generation and cell cycle arrest. PLoS One. 2014 Oct 16;9(10):e110003. [3]. Cho KW, et al. Dietary naringenin increases hepatic peroxisome proliferators-activated receptor α proteinexpression and decreases plasma triglyceride and adiposity in rats. Eur J Nutr. 2011 Mar;50(2):81-8. [4]. Mulvihill EE, et al. Naringenin prevents dyslipidemia, apolipoprotein B overproduction, and hyperinsulinemia in LDLreceptor-null mice with diet-induced insulin resistance. Diabetes. 2009 Oct;58(10):2198-210. [5]. Chen S, et al. Naringenin inhibits TNF-α induced VSMC proliferation and migration via induction of HO-1. Food Chem Toxicol. 2012 Sep;50(9):3025-31. |